
科研进展
《Plant Physiology》在线发表万建民院士团队“OsVPS34-generated PI3P recruits GPA5/Rab5a to regulate post-Golgi glutelin trafficking in rice endosperm”
发布人: 发布日期: 2026-03-26 浏览次数:
水稻是全球半数以上人口的主粮,其种子储存蛋白占籽粒干重的8-10%,是人类膳食植物蛋白的重要来源。其中谷蛋白占总储存蛋白的60-80%,其合成与分选过程直接决定稻米的营养品质、加工特性和致敏性。谷蛋白前体在内质网合成后,经高尔基体加工,通过致密囊泡(DV)转运至蛋白储存液泡(PBII),被切割为成熟的酸性和碱性亚基后沉积。此前研究已鉴定到多个调控谷蛋白转运的蛋白因子,但磷脂类信号分子在该过程中的功能与调控机制长期未被阐明,而III类磷脂酰肌醇3-激酶(PI3K)核心亚基VPS34的纯合突变致死,更是极大限制了其在种子发育中的功能解析。
2026年3月,国际植物学权威期刊《Plant Physiology》在线发表了南京农业大学/中国农业科学院作物科学研究所万建民院士、董慧教授、王益华教授团队题为“OsVPS34-generated PI3P recruits GPA5/Rab5a to regulate post-Golgi glutelin trafficking in rice endosperm”的研究论文。该研究利用筛选到的谷蛋白前体积累弱突变体gpa14,结合图位克隆、脂质组学、细胞生物学、生物化学等多维度技术,首次明确了磷脂酰肌醇3-磷酸(PI3P)在谷蛋白转运中的核心作用,揭示了OsVPS34介导的PI3P合成通过招募调控因子GPA5/Rab5a,促进致密囊泡与蛋白储存液泡融合,保障谷蛋白正确沉积的分子机制,为稻米蛋白品质的分子改良提供了关键理论靶点和基因资源。
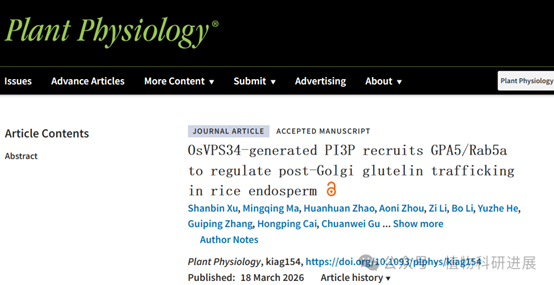
1. gpa14突变体表现出谷蛋白前体过度积累的粉质胚乳表型
研究从组织培养诱变的粳稻品种宁粳4号突变体库中筛选获得谷蛋白前体积累突变体gpa14,其成熟种子表现为粉质皱缩表型,扫描电镜显示突变体胚乳淀粉颗粒呈球形、排列松散。SDS-PAGE和免疫印迹分析显示,gpa14中57 kDa谷蛋白前体大量积累,而成熟谷蛋白酸性/碱性亚基、26 kDa α-球蛋白、醇溶蛋白的含量均显著降低。内质网分子伴侣BiP1和PDI1-1的含量在突变体与野生型中无显著差异,说明谷蛋白前体积累并非内质网输出缺陷导致,而是高尔基体后的转运过程出现异常。 细胞学观察证实了转运缺陷:12 DAF(授粉后天数)胚乳的免疫荧光标记显示,野生型中谷蛋白和α-球蛋白共同定位于PBII中,而gpa14中部分谷蛋白和α-球蛋白被错误分选到胞外形成副壁体(PMB)结构,PBII的体积显著小于野生型,仅储存醇溶蛋白的PBI结构未受影响。透射电镜观察进一步发现,gpa14中高尔基体出芽形成的致密囊泡(DV)无法正常转运至PBII,大量堆积在细胞膜附近并被分泌到胞外,形成电子致密的PMB结构。
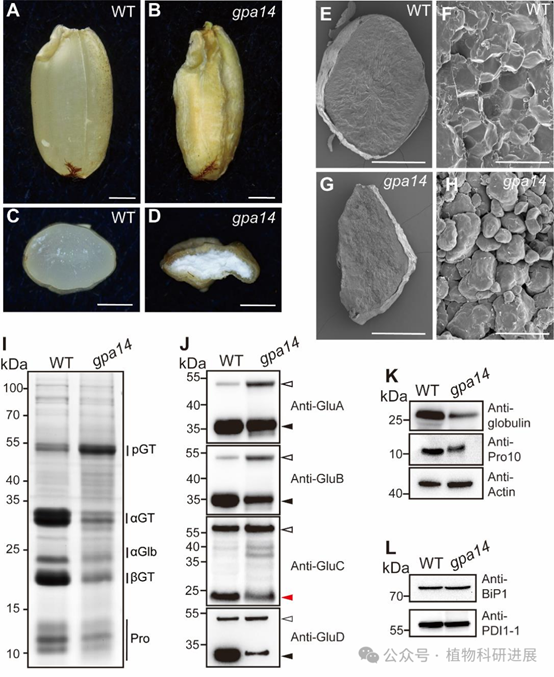
图1 gpa14突变体的表型鉴定
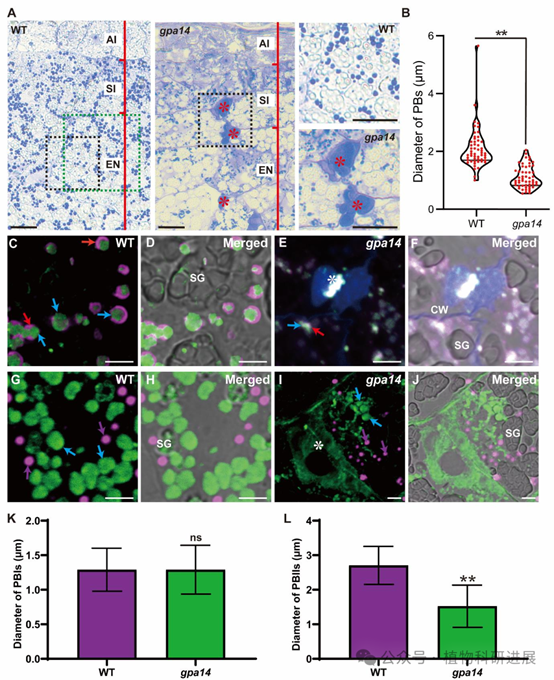
图2 gpa14突变体中谷蛋白错误分选形成副壁体
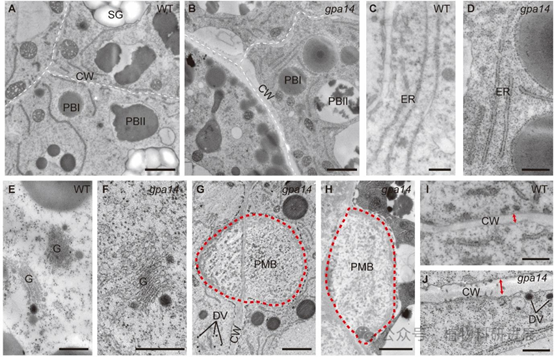
图3 gpa14突变体胚乳亚细胞结构的透射电镜观察
2. 图位克隆证实GPA14编码III类PI3K核心亚基OsVPS34
利用gpa14杂合株与籼稻品种N22构建的F2分离群体,研究将目标基因初定位到5号染色体2.1-5.5 Mb的区间内。对野生型和突变体进行全基因组重测序分析发现,该区间内Os05g0180600基因的第四外显子存在一个单胸腺嘧啶插入,导致移码突变,翻译提前终止,缺失了C端的PI3Ka和PI3_PI4_激酶功能结构域,该基因编码III类磷脂酰肌醇3-激酶的催化亚基OsVPS34。 遗传互补实验验证了基因功能:在gpa14突变体背景下过表达全长OsVPS34,可完全恢复突变体的粉质胚乳表型,谷蛋白前体积累、PBII结构缺陷和PMB结构均消失,证实OsVPS34是调控谷蛋白转运的目标基因。进化分析显示OsVPS34在单双子叶植物中高度保守,其在水稻各组织中组成型表达,在发育胚乳中持续高表达,与谷蛋白合成时期高度吻合;亚细胞定位显示OsVPS34定位于细胞质和细胞核。
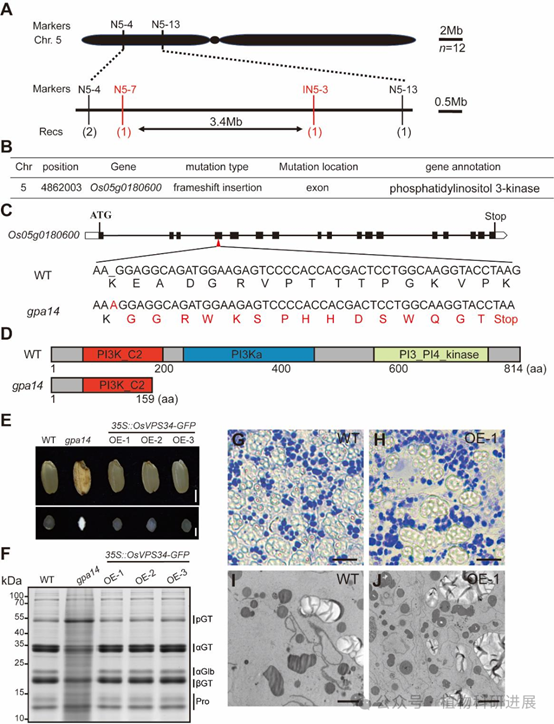
图4 GPA14的图位克隆与遗传互补验证
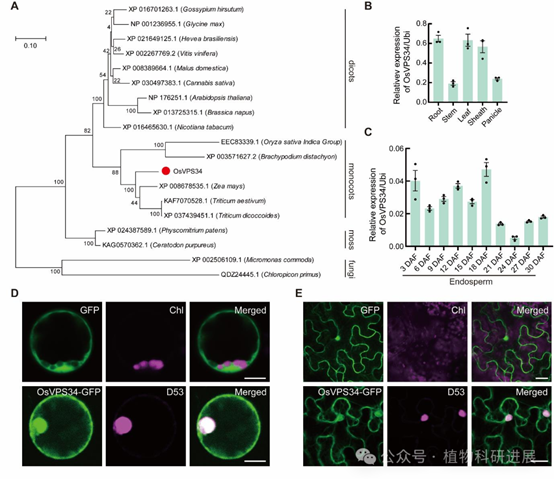
图5 OsVPS34的进化、表达模式与亚细胞定位
3. OsVPS34作为PI3K复合体组分催化PI3P合成,且PI3P定位于谷蛋白转运的关键细胞器
为解析OsVPS34的作用机制,研究通过免疫共沉淀联合质谱(IP-MS)鉴定到OsVPS34的互作蛋白包括PI3K复合体的核心亚基OsVPS15、OsATG6b、OsVPS38和OsATG14,说明水稻中存在保守的两类PI3K复合体:调控自噬的PI3K复合体I(含ATG14)和调控囊泡转运的PI3K复合体II(含VPS38)。进一步通过酵母双杂交、荧光素酶互补(LCI)、Co-IP、双分子荧光互补(BiFC)实验证实,OsVPS34与OsATG6b存在最强的直接互作,且OsVPS34-OsATG6b复合体共定位于反式高尔基体网络(TGN)和前液泡区室(PVC),与谷蛋白转运的关键细胞器定位一致。 PI3K复合体的核心功能是催化合成磷脂酰肌醇3-磷酸(PI3P),检测显示gpa14发育胚乳中PI3P的含量较野生型降低60%以上,免疫荧光标记显示突变体中PI3P的点状信号数量仅为野生型的20%。利用PI3P特异性报告系统FYVE-GFP的共定位分析显示,PI3P主要富集于TGN和PVC,皮尔逊相关系数分别达0.793和0.827;免疫金标记进一步证实PI3P与谷蛋白共定位于致密囊泡(DV)和PBII中,直接证明PI3P参与谷蛋白的转运过程。
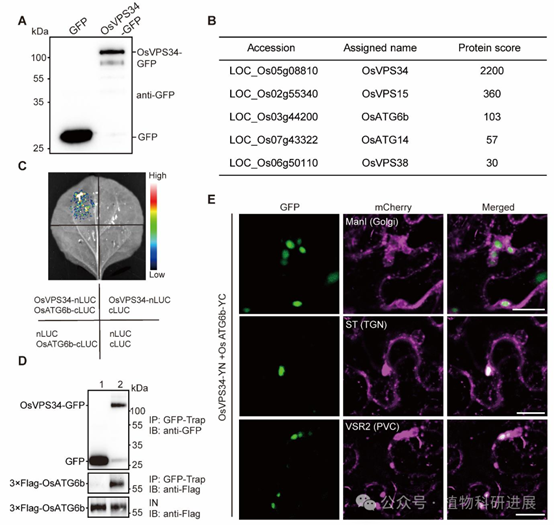
图6 水稻PI3K复合体的互作验证
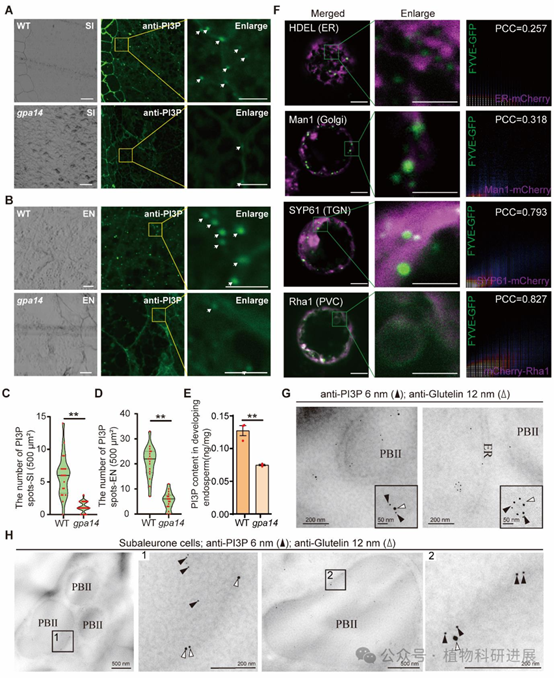
图7 水稻胚乳中PI3P的含量与亚细胞定位
4. PI3P通过招募GPA5和Rab5a到膜系统调控谷蛋白转运
GPA5是已报道的Rab5a效应子,通过N端PX结构域结合PI3P,调控DV与PBII的融合过程。为解析PI3P的下游调控机制,研究通过细胞组分分离实验发现:野生型中GPA5主要定位于膜组分(P100),而gpa14突变体中膜结合型GPA5的含量显著降低,更多分布于细胞质组分(S100)。利用PI3K抑制剂LY294002处理Rab5a-GFP转基因株系,同样发现Rab5a和GPA5的膜结合比例显著下降。 进一步构建地塞米松(DEX)诱导的OsVPS34 RNAi株系,DEX处理后OsVPS34表达量显著下调,PI3P含量降低,此时Rab5a和GPA5与PVC标记Rha1的共定位程度较对照组下降40%以上,证实PI3P是维持二者膜定位、保障其功能的关键信号。
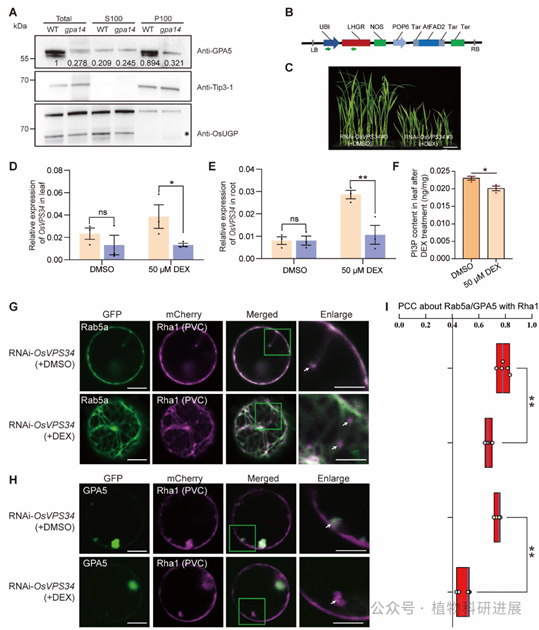
图8 PI3K复合体功能破坏后Rab5a与GPA5的定位异常
5. OsVPS34调控谷蛋白转运的分子工作模型
基于上述结果,研究提出了完整的调控模型:野生型水稻胚乳中,OsVPS34作为PI3K复合体的核心催化亚基,在TGN出芽形成的致密囊泡(DV)和PBII膜上催化合成PI3P;PI3P作为膜信号分子,招募效应子GPA5和小G蛋白Rab5a到膜上,后续GPA5通过与CORVET栓系复合体、SNARE融合复合体互作,介导DV与PBII的膜融合,将谷蛋白前体转运至PBII中完成切割和沉积;而gpa14突变体中OsVPS34功能缺失,PI3P合成受阻,GPA5和Rab5a无法正确定位到膜系统,导致DV不能正常转运至PBII,未被转运的谷蛋白被错误分泌到胞外形成PMB结构,最终表现为谷蛋白前体积累、胚乳粉质皱缩的表型。
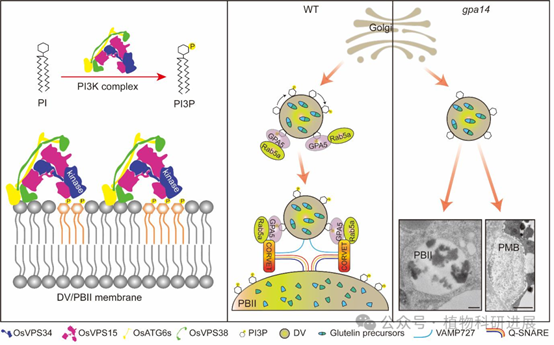
图9 OsVPS34调控水稻胚乳谷蛋白高尔基体后转运的工作模型
本研究克服了PI3K核心亚基纯合突变致死的研究瓶颈,利用仅表现胚乳表型的弱突变体gpa14,首次直接证明了磷脂信号分子PI3P在水稻谷蛋白转运过程中的核心功能,完善了种子储存蛋白分选的调控网络,同时解析了水稻PI3K复合体的组成与亚细胞定位,为后续PI3K通路在植物发育和逆境响应中的功能研究提供了新方向。 该研究挖掘的OsVPS34、GPA5等关键基因为稻米蛋白品质的分子改良提供了直接靶点:通过编辑调控该通路的活性,可定向优化谷蛋白的含量与组成,既可以提升谷蛋白含量培育高营养稻米,也可以降低谷蛋白含量培育适合肾病患者食用的特殊功能性稻米;同时该调控模块的保守性也可拓展到小麦、玉米等其他禾本科作物的品质改良中。后续可进一步解析PI3P调控GPA5蛋白稳定性的分子机制,挖掘OsVPS34的优异等位变异,开发功能分子标记,加速高产优质水稻新品种的培育进程。
---原文链接--- https://doi.org/10.1093/plphys/kiag154
- 上一篇: 《JIPB》上发表盖钧镒院士团队“Constitution and function of intergenic genome regions in cultivated versus annual wild soybean in China”
- 下一篇: 《International Journal of Applied Earth Observation and Geoinformation》发表智慧农业创新团队“Accurate estimation of grain number per panicle in winter wheat by synergistic use of UAV imagery and meteorological data “